今天seo就给大家介绍酮体代谢及酮体的多种生理功能的文章,看完这篇文章相信你对酮体代谢及酮体的多种生理功能介绍更加了解!
酮体代谢及酮体的多种生理功能
脂解产生的乙酰辅酶A除了进入三羧酸循环氧化之外,还可以在肝脏中合成酮体(ketone body)。酮体指乙酰乙酸、-羟基丁酸和丙酮这三种物质。
酮体主要是在缺乏葡萄糖时作为肝外组织的能源物质。心脏和肾上腺皮质主要以酮体作燃料,骨骼肌也可利用酮体,脑在平时利用葡萄糖,但在饥饿时会将酮体作为主要燃料,将珍贵的葡萄糖留给红细胞。在某些病理条件下(如糖尿病等),也会生成大量酮体。
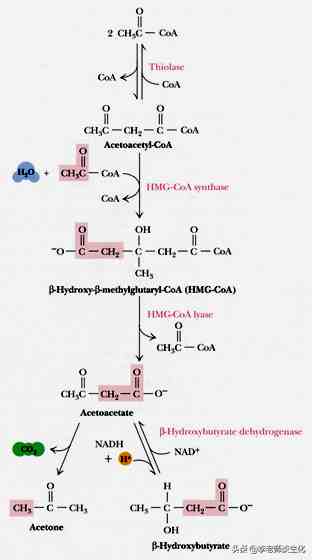
酮体的合成首先是两个乙酰辅酶A缩合生成乙酰乙酰辅酶A。这个反应可以看作-氧化最后一步的逆转,但所用的酶是另外一个酶,线粒体乙酰乙酰辅酶A硫解酶(mThiolase),或称乙酰辅酶A乙酰转移酶(acetyl-CoA acetyltransferase),由ACAT1基因编码。这样有利于两个过程的分别调控。
酮体的合成
乙酰乙酰辅酶A再与第三个乙酰辅酶A缩合,就生成-羟基--甲基戊二酰辅酶A(HMG-CoA)。催化的酶是HMG辅酶A合酶,由HMGCS2基因编码。这两步反应与胆固醇合成的起始反应相同,但后者在细胞质中进行,所以负责催化的是胞浆同工酶(ACAT2和HMGCS1)。
HMG-CoA裂解酶(HMGCL)将其裂解,生成乙酰-CoA和乙酰乙酸(AcAc或AA)。这是酮体合成的限速步骤。另外,HMGCL还参与亮氨酸的碳架氧化。
D--羟丁酸脱氢酶(BDH1)催化乙酰乙酸的还原,生成羟丁酸(BHB或OHB),反应可逆,不催化L-型底物。乙酰乙酸可以自发或由乙酰乙酸脱羧酶催化脱羧,生成丙酮。
酮体可通过自由扩散或由单羧酸转运蛋白1(MCT1)或MCT2转运出线粒体,二者分别由SLC16A1和SLC16A7基因编码。
BHB比AA更稳定,所以是血液中最主要的酮体。人体BHB的基础血清水平在M范围,但禁食12-16小时后会上升到几百M,禁食2天后达到1-2 mM,长期饥饿可达到6-8 mM(Trends Endocrinol Metab. 2014 Jan;25(1):42-52.)。
酮体被肝外组织吸收后,有两条代谢途径。一条是进入线粒体氧化分解,称为氧化代谢命运,这是其主要代谢方式;另一条是在细胞质中参加脂合成,称为非氧化代谢命运。
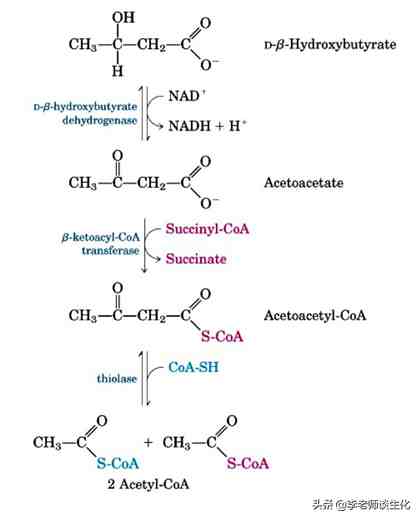
BHB可由BDH1重新氧化生成乙酰乙酸,然后生成乙酰乙酰辅酶A,再通过-氧化和三羧酸循环提供能量。生成乙酰乙酰辅酶A是酮体利用的限速步骤,线粒体中所用的酶是3-酮脂酰辅酶A转移酶1(OXCT1),也称为琥珀酰辅酶A:酮脂酰辅酶A转移酶(SCOT)。
酮体的氧化代谢
这是一个辅酶A的交换反应,好处在于不需要消耗ATP,由三羧酸循环提供琥珀酰辅酶A。但其平衡有利于逆反应,需要靠酮体的不断消耗来推动。SCOT在肝脏以外的所有线粒体中均有表达,这也是肝脏能不断输出酮体的原因。
细胞质中的反应由乙酰乙酰辅酶A合酶催化,然后乙酰乙酰辅酶A可被胞质HMGCS1催化进入胆固醇合成,或被ACAT2催化进入脂肪酸合成。
丙酮代谢比较复杂,先被单加氧酶催化羟化,然后可生成丙酮酸或乳酸、甲酸、乙酸等。大部分丙酮异生成糖,这是脂肪酸转化为糖的一个可能途径,但平时流量极低,只有在酮症状态下才有意义。
酮体的主要生理意义是作为禁食能源。这在进化上是很保守的,许多细菌合成OHB的聚合物来存储能量。这种反应可用于生产生物降解塑料,如聚羟基丁酸(PHB)及其改进产品PHBHV。它们可降解、无毒,由可再生资源通过微生物发酵生产。已有多种此类商品上市,用于食品、药品等领域。例如有人用PHBHV制成生物相容性纳米载体,用于靶向大肠癌的5-FU递送(Drug Deliv. 2019 Dec;26(1):318-327.)。
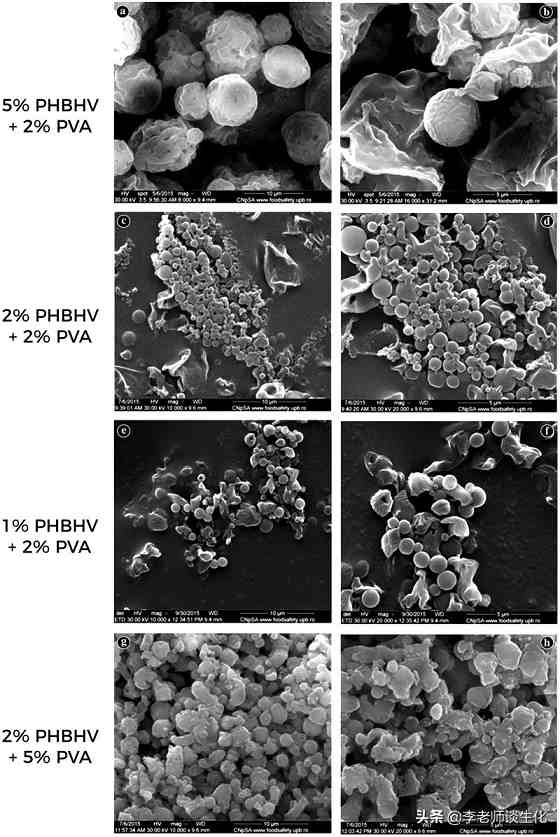
PHBHV纳米载体颗粒SEM显微照片,引自Drug Deliv. 2019 Dec;26(1):318-327.
酮体代谢与糖脂代谢密切相关。它与细胞质和线粒体中的代谢途径是整合在一起的。当肝脏中有足够的磷酸二羟丙酮时,就会有充足的磷酸甘油,大多数脂肪酸会用来合成甘油三酯。当脂肪酸进入线粒体后,如果对ATP的需求不高,就会用来合成酮体。
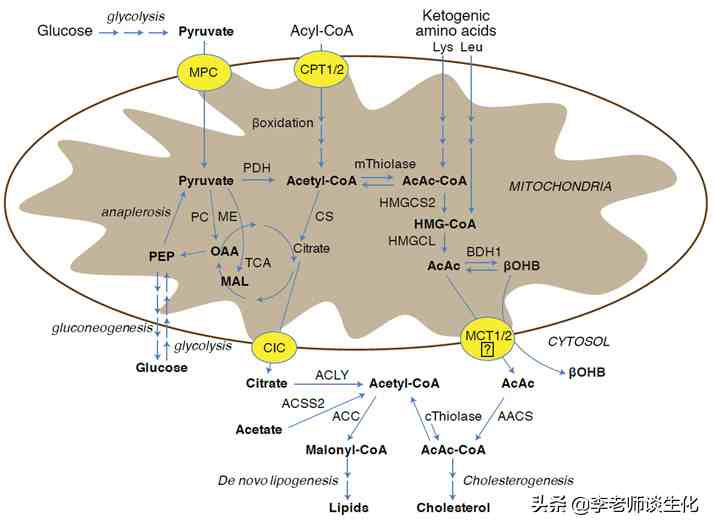
酮体代谢与细胞质代谢途径的整合,引自Cell Metab. 2017 Feb 7;25(2):262-284.
酮体合成的关键酶HMGCS2主要受FOXA2、PPAR和FGF21的转录调节,以及琥珀酰化和乙酰化/ SIRT3脱乙酰化的翻译后调控。酮体降解的关键酶OXCT1则主要受琥珀酰辅酶A调控。
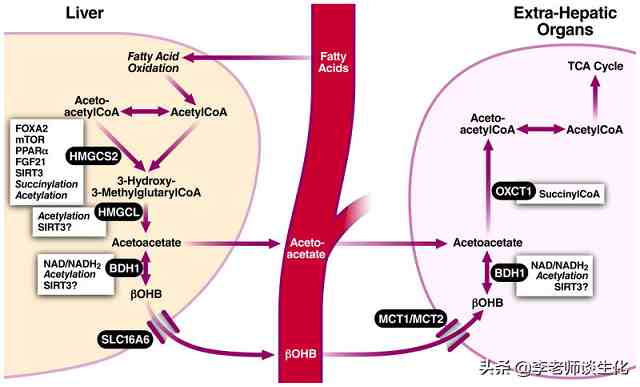
酮体代谢调控,引自Trends Endocrinol Metab. 2014 Jan;25(1):42-52.
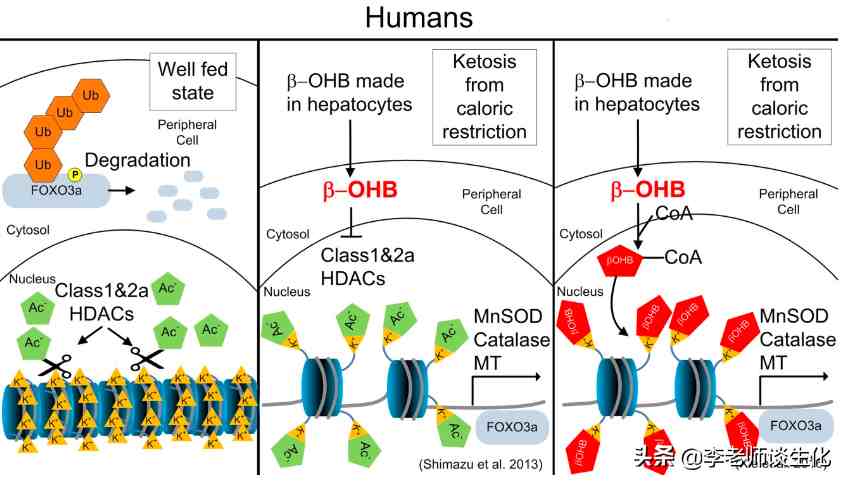
很多代谢物都有信号传导作用,酮体也不例外。BHB作为最主要的循环酮体,当仁不让地成为了一个信号分子,为酮体代言。BHB至少有两种G蛋白偶联受体(GPCR),包括HCAR2(GPR109A)和FFAR3(GPR41),可以减少脂肪分解,减少交感神经紧张,降低代谢率。这与禁食状态相一致。
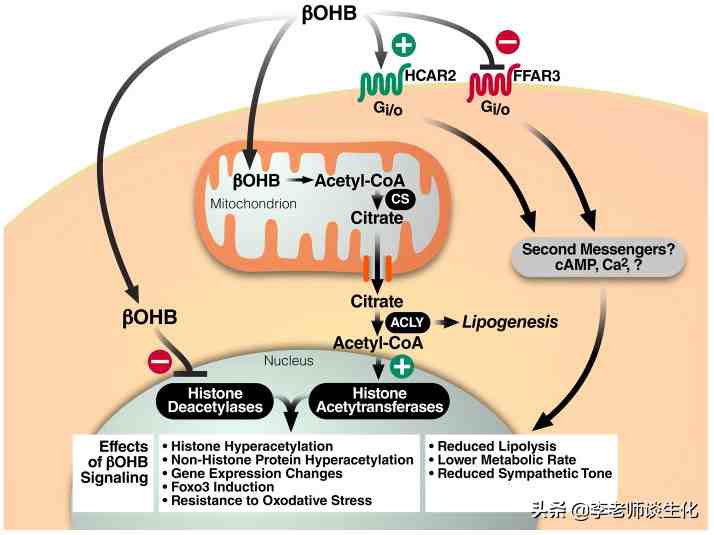
-羟丁酸介导的信号转导,引自Trends Endocrinol Metab. 2014 Jan;25(1):42-52.
BHB具有表观遗传调控作用。它可抑制组蛋白脱乙酰基酶(HDAC)活性,促进一些抗氧化基因的表达,从而降低细胞氧化应激水平。在长期禁食条件下,BHB还可以使组蛋白中赖氨酸残基-羟基丁酰化。这是表观遗传调控的一种新形式,可以改变多种基因表达,例如转录共激活因子PGC-1,从而调控IRS2和CPT1A等。
此外,酮体还与炎症、肿瘤、神经保护以及寿命调控等多种事件相关。